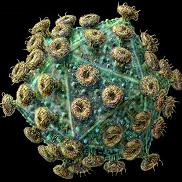
| باکتریوفاژها ویروسهایی هستند که از باکتریهای مختلف به عنوان سلول میزبان استفاده میکنند، لذا انگل یا پارازیت داخل سلول باکتریایی هستند. هر باکتریوفاژ در یک باکتری اختصاصی و مناسب خود رشد میکند. در طبیعت انواع مختلف باکتری و متناسب با آنها انواع مختلف فاژها وجود دارد. |
دید کلی
میزبان
ویروسها عبارت است از گونههایی از میزبان که ویروس بتواند آنها را آلوده سازد. ویروسها فقط در درون گونههای خاصی تکثیر پیدا میکنند و از اینرو آنها را به سه گروه اصلی به نام
ویروسهای جانوری ،
ویروسهای گیاهی و
ویروسهای باکتریایی یا باکتریوفاژها تقسیم میکنند. در هر رده هر نوع ویروس معمولا سلولهای گونه خاصی را میتواند آلوده سازد. میزبانهای خاص یک ویروس بوسیله نیاز لازم برای اتصال اختصاصی ویروس به سلول میزبان و در دسترس بودن عوامل میزبانی ضروری برای
تکثیر ویروسها تعیین میگردد.
برای آلوده شدن میزبان بوسیله ویروس سطح خارجی ویروس باید با پذیرندههای اختصاصی سطح سلول میزبان واکنش نشان دهد. در برخی از باکتریوفاژها نقطه پذیرنده یک نوع ماده شیمیایی دیواره سلول میزبان است. باکتریوفاژها پس از تکثیر در داخل باکتریها ، آنها را لیز میکنند. بنابراین به نام
باکتریوفاژ یا به اختصار
فاژ (خورنده باکتری) معروفند. توضیح اینکه هر باکتریوفاژ در یک
باکتری اختصاصی و مناسب خود رشد میکند. در طبیعت انواع مختلف باکتری و متناسب با آنها انواع مختلف فاژها وجود دارد.
تاریخچه کشف باکتریوفاژها
در سالهای 1915 و 1917 میلادی دو دانشمند به نامهای توورت و هرله در ضمن آزمایشات خود بطور اتفاقی به وجود این فاژها پی بردند. این دانشمندان ضمن رشد باکتریهای مختلف در محیطهای کشت مایع (مثل آبگوشت) متوجه مردن و یا لیز شدن خودبهخود باکتریها شدند. این دو دانشمند پس از مطالعات کوتاه مدت و با صاف نمودن این محیطهای حاوی فاژ توسط
فیلترهای باکتریولوژی وجود این باکتریوفاژها را اثبات کردند.
مختصری از ویروس بدانیم
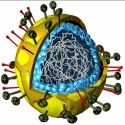
ویروسها کوچکترین
میکروارگانیسمها هستند که با
میکروسکوپ نوری قابل رویت نیستند. اندازه آنها بین 300 - 20 نانومتر متفاوت است و تنها با
میکروسکوپ الکترونی قابل مشاهدهاند. ویروسها پارازیتهای داخل سلولیاند که برای تکثیر به یک سلول زنده (
یوکاریوت یا
پروکاریوت ،
سلول گیاهی یا
پروتوزوئرها و یا
قارچها و ... ) نیاز دارند. آنها فقط قادرند در سیستمهای زنده تکثیر پیدا کنند و در محیطهای کشت مصنوعی تکثیر نمیشوند. کلیه ویروسها فقط یک نوع
اسید نوکلئیک دارند، یا
DNA دارند و یا فقط
RNA دارند. هر دو نوع اسید نوکلئیک در یک ویروس دیده نمیشود. به استثنای
ویروسهای سرطانزا.
ویروسها پوشش پروتئینی محافظ اسید نوکلئیک به نام
کپسید دارند که از واحدهایی به نام
کپسومر تشکیل یافته است. برخی ویروسها حاوی غلاف هستند که آن را از غشای میزبان گرفتهاند. در هیچ یک از ویروسها سیستمهای انرژیزا نظیر
میتوکندری و
ریبوزوم وجود ندارد، لذا اجبارا پارازیت داخل سلولیاند. ویروسها به درجات حرارت بالا حساس بوده و خاصیت بیماریزای خود را از دست میدهند، ولی بالعکس به درجات حرارت پایین صفر (70- تا 196- درجه سانتیگراد) مقاومند و خاصیت بیماریزایی خود را حفظ میکنند. ویروسها به
آنتی بیوتیکها حساس نیستند. در برخی از موارد یاد شده استثناهایی وجود دارد، مثلا
ویروس هپاتیت B درجه حرارت جوش را نیز تحمل میکند.
تقسیمبندی فاژها
باکتریوفاژها را امروزه برحسب نوع جنس آنها در گروههای مختلف تقسیمبندی میکنند. باکتریوفاژها نیز نظیر سایر
ویروسها از یک
اسید نوکلئیک و
کپسید تشکیل شدهاند، ولی در بعضی از فاژها قسمتهای مختلف دیگری مثل
دم ،
غلاف روی دم ،
پایه انتهایی و
رشتههای خار مانند نیز دیده میشود. امروزه باکتریوفاژها را برحسب نوع و شکل در 6 گروه تقسیمبندی میکنند.
فاژهای دمدار با غلاف کوتاه شونده
این دسته از فاژها شکل تیپیک فاژها را تشکیل میدهند و از سه قسمت کپسید (بهعنوان سر) ، دم و پایهای که در انتهای دم قرار دارد تشکیل شدهاند. کپسید آنها 8 وجهی (ایکوزاهورال یا اکتاهورال) میباشد. داخل کپسید اسید نوکلئیک
DNA دورشتهای قرار دارد. مثالهای این گروه از فاژها عبارتند از: فاژهای T
6 ، T
4، T
2 برای
باکتری Ecoli و یا فاژهای P
1 و P
2 برای
باکتری سالمونلا .
این دسته از فاژها در زمان حمله و نفوذ به درون
باکتری مورد نظر با زواید خارمانندی که در انتهای دم خود دارند، بر روی باکتریها میچسبند. سپس غلاف روی دم آنها منقبض میشود (جمع میشود) و مثل یک سرنگ عمل میکند، قسمت دم وارد دیواره باکتری شده و DNA موجود در داخل کپسید از داخل سوراخ وسط کانال دم عبور کرده و به داخل
سیتوپلاسم باکتری وارد میشود. بقیه قسمتهای ویروس در بیرون از سلول باکتری باقی میماند.
فاژهای دمدار با دم دراز بدون غلاف
این گروه دارای یک کپسید و یک دم دراز بدون غلاف هستند. شکل کپسید شبیه کپسید گروه اول است. در داخل آن اسید نوکلئیک DNA دو رشتهای وجود دارد. نحوه ورود اسید نوکلئیک این فاژها در زمان آلوده کردن باکتریها هنوز مشخص نشده است. این گروه از فاژها در طبیعت بسیار فراوانند و تقریبا برای تمامی
انواع باکتریها از این فاژها مشخص شده است: مثل T
5 ، T
1 و فاژ لاندا.
فاژهای بادم کوتاه و بدون غلاف
کپسید این گروه نیز شبیه دو گروه اول بوده ولی دم آنها بسیار کوتاه است. برخی از فاژهای موجود در این گروه در انتهای خود فقط دارای یک زایده کوچک هستند، یعنی طول دم در این فاژها از طول کپسید به مراتب کوتاهتر است. اسید نوکلئیک آنها DNA دورشتهای است. فاژ T
7 ، T
3 برای Ecoli و فاژ P
22 برای باکتری سالمونلا در این گروه هستند.
باکتریوفاژهای بیدم با کپسومر بزرگ
در این فاژها بطور کلی دم و زواید آن وجود ندارد و تنها از کپسید تشکیل یافتهاند. اسید نوکلئیک آن DNA تکرشتهای است و این ویروسها تقریبا کوچکند به کوچکی
ویروس فلج اطفال یعنی 25 نانومتر. کپسومرهای آن بزرگند. مثال آن M
20 و S
13 برای باکتری Ecoli است.
باکتریوفاژهای بیدم با کپسومر کوچک
کپسید این فاژها ، هم 8 وجهی بوده، هم کوچک است و ساختمان خیلی ساده دارد که از 92 کپسومر تشکیل شده است که یکی از این کپسومرها بزرگتر از بقیه بوده و در یک گوشه کپسید میباشد و احتمال میدهند که این کپسومر بزرگ نقش چسبیدن فاژ را به باکتری بازی میکند. این گروه از فاژها دو ویژگی مهم دارند اول اینکه اسید نوکلئیک داخل کپسید آنها از
RNA تکرشتهای تشکیل شدهاست و دوم اینکه این دسته از فاژها فقط باکتریهای مذکر یا دنور را آلوده میکنند و به کنارههای پیلوسهای جنسی میچسبند. مانند فاژهای FR ، MS
2 ، F
2 برای Ecoli.
فاژهای میله مانند یا فیلامانتوز
برخلاف بقیه فاژها این دسته از فاژها ساختمانی شبیه به کپسید ندارند و به صورت میله مانند دیده میشوند. در داخل ساختمان این گروه از فاژها
اسید نوکلئیک DNA تکرشتهای وجود دارد. این گروه نیز همانند گروه پنجم فقط باکتریهای مذکر را آلوده میکنند و به قسمت نوک و انتهای پیلوسهای جنسی میچسبند، مثل فاژ fd ، M
13 ، F
1 برای Ecoli.
سیکلهای حیاتی
فاژها در حالت کلی دو سیکل حیاتی
لیتیک و
لیزوژنیک دارند.
سیکل حیاتی لیتیک
در سیکل حیاتی لیتیک فاژها پس از ورود به داخل باکتری حساس بهخود تکثیر یافته و
همانندسازی میکنند و باکتری را در نهایت لیز میکنند. لیز شدن باکتری بدین صورت انجام میگیرد که از طرف
ژنوم فاژیک پروتئین لیزکننده سنتز میشود و این پروتئین دیواره باکتری را از داخل از بین میبرد و دیواره را متلاشی میکند. لذا فاژهای تشکیل یافته جدید بیرون میریزند که در این حالت به آنها
progeny phage گویند که در تماس با باکتری جدید به آن میچسبند. و این سیکل دوباره تکرار میشود.
سیکل حیاتی لیزوژنیک
بعضی از فاژها بصورت temprator زندگی میکنند و بعد از ورود به باکتری رشد و تکثیر نمییابند و باکتری را لیز نمیکنند. بدین صورت که ژنوم فاژ وارد شده به داخل باکتری به
ژنوم باکتری چسبیده و ادغام میشود و همراه ژنوم باکتری با Replication
کروموزوم باکتری تقسیم شده و به نسلهای بعدی نیز منتقل میشود. اسید نوکلئیک فاژ که با این مکانیزم به نسلهای بعدی منتقل میشود، به نام پروفاژ معروف است و به این عمل نیز عمل لیزوژنیک گویند.
باکتریهایی که در ژنوم خود یک ژنوم فاژ را حمل میکنند، معمولا به آلودگیهای بعدی توسط فاژها مقاومند. این عمل لیزوژنی به بعضی از باکتریها خواص پاتوژنی میدهد و یا در عدهای دیگر مثلا در
باسیل دیفتری و
استرپتوکوک اعمال سنتز
توکسینهای مختلف باکتری توسط پروفاژ موجود در
کروموزوم آن انجام میشود. این پروفاژ موجود در کروموزوم باکتری به علل مختلف میتواند از کروموزوم جدا شده و وارد مرحله لیتیک شود، در این صورت تکثیر یافته و باکتری را لیز میکند.
مراحل ورود و تکثیر فاژها
مرحله تماس و یا جذب فاژ بر روی سلول باکتری
در فاژهای دمدار زواید دم در چسبیدن فاژ بر روی سلول باکتری نقش دارند. البته فعالیت
لیزوزومها برای بوجود آوردن سوراخ یا نقطهای در روی دیواره باکتری نیز ضروری است. در فاژهای بیدم بعضی از کپسومرهای موجود در روی کپسید فاژ این عمل را انجام میدهند. برای انجام عمل جذب در محیط حاوی باکتری و فاژ اختصاصی آن وجود املاحی نظیر
کلرید سدیم یا کلرید کلسیم الزامی است.
مرحله ورود فاژ به داخل باکتری
در فاژهای گروه اول اسید نوکلئیک داخل کپسید با کوتاه شدن غلاف روی دم و ورود قسمتی از دم به دیواره باکتری ، از راه کانال دم وارد باکتری میشود. برخلاف ویروسهای انسانی و حیوانی فقط اسید نوکلئیک وارد شده و بقیه زواید در بیرون از
سلول باکتری باقی میمانند. یعنی مرحله uncoating در تکثیر سایر ویروسها یعنی خروج اسید نوکلئیک از کپسید در فاژها ضمن ورود آنها به داخل باکتری صورت میگیرد.
بیوسنتز
در این مرحله اسید نوکلئیک فاژ کلیه اعمال سنتز خود را برعهده گرفته و این اعمال را در داخل باکتری کنترل میکند. اول mRNA و پروتئینهای Early (اولیه) را سنتز میکند که اینها خاصیت آنزیماتیکی دارند و برای ساختهشدن اسید نوکلئیک جدید بکار میروند و پس از آن mRNA و پروتئینهای ثانویه را سنتز میکند که در ساختمان فاژهای جدید بکار خواهند رفت.
رسیدگی کامل
در این مرحله اسید نوکلئیک تولیدشده به حالت فشرده در آمده و توسط
کپسومرها احاطه میشود و بالاخره کپسید تشکیل پیدا کرده و در داخل آن قرار میگیرد. همزمان با این اعمال دم فاژها نیز ساخته میشود و در روی کپسید به محلهای مخصوص به خودشان میچسبند و آرام آرام فاژهای جدید تشکل مییابند.
برگرفته از : دانشنامه رشد



 به نام خدا :
به نام خدا :